進化学実習 2022 牧野研
7. 統計モデリング2: 一般化線形モデル
統計モデリングのお品書き

久保先生の"緑本"こと
「データ解析のための統計モデリング入門」
をベースに回帰分析の概要を紹介。
回帰のキモは線ではなく分布。
Data Science Hill Climb 2021 (東京海上) での講義 (~6時間) の演習無し抜粋バージョン (~2時間 x 2回)。
何でもかんでも直線あてはめではよろしくない
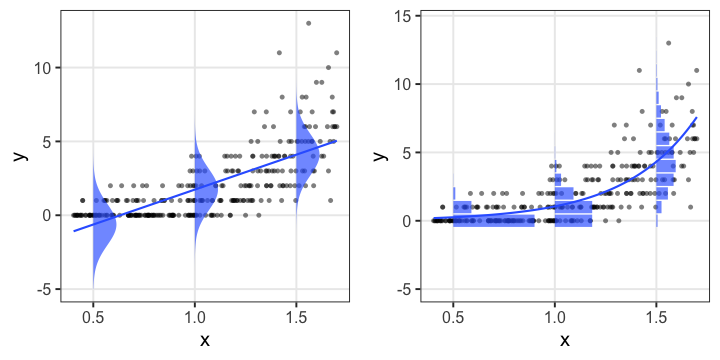
- 観察データは常に正の値なのに予測が負に突入してない?
- 縦軸は整数。しかものばらつきが横軸に応じて変化?
- データに合わせた統計モデルを使うとマシ
ここまで見てきた統計モデル
確率変数$X$はパラメータ$\theta$の確率分布$f$に“従う”: $X \sim f(\theta) $
e.g., ある植物が作る種の数$X$は平均値$\lambda$のポアソン分布に従う:
これを一般化線形モデル(GLM)として見ることもできる。
一般化線形モデル(GLM)として記述してみる
個体$i$の種子数$y_i$は平均値$\lambda_i$のポアソン分布に従う。
平均値$\lambda_i$は他のデータによらず$\beta_0$で一定。
種子数をY軸にして、式を2つに分けただけ…?
説明変数を含むモデルを見ればご利益が分かるかも。
説明変数が1つある一般化線形モデル
個体$i$の種子数$y_i$は平均値$\lambda_i$のポアソン分布に従う。
平均値の対数$\log(\lambda_i)$はその個体の大きさ$x_i$に比例する。

この場合は単回帰。説明変数が複数あると重回帰。
複数の説明変数を同時に扱う重回帰
\[\begin{split} y_i &\sim \text{Poisson}(\lambda_i) \\ \log(\lambda_i) &= \beta_0 + \beta_1 x_{1i} + \beta_2 x_{2i} + \ldots \end{split}\]
気温も湿度も高いほどビールが売れる、とか
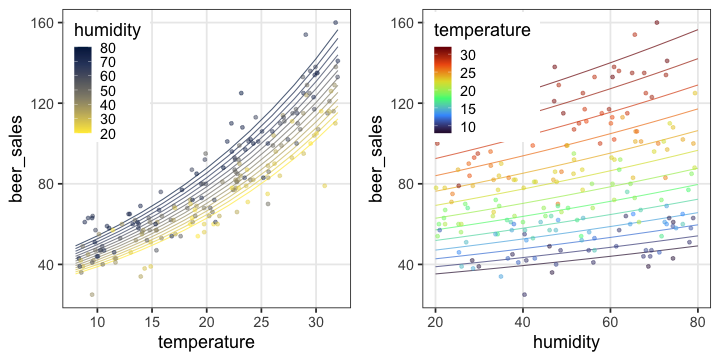
今度は確率分布とリンク関数を変えてみよう。
ロジスティック回帰
- 確率分布: 二項分布
- リンク関数: $\text{logit}(p) = \log \frac {p} {1 - p}$
何かの成否に対する何かの因子の影響、とか
客10人中$y_i$人がビールを注文。
その日$i$の気温$x_i$によって割合が変化。
\[\begin{split} y_i &\sim \text{Binomial}(n,~p_i) \\ \text{logit}(p_i) &= \beta_0 + \beta_1 x_i \\ p_i &= \frac 1 {1 + e^{-(\beta_0 + \beta_1 x_i)}} \end{split}\]
ロジスティック関数↑
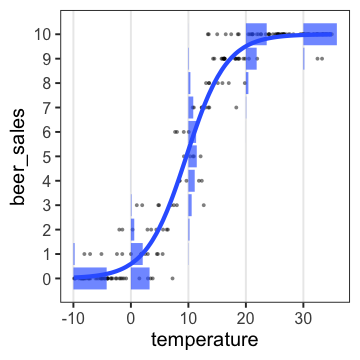
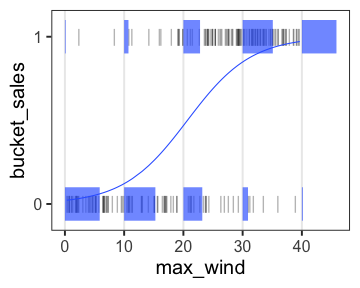
ロジスティック回帰 (狭義)
- 確率分布: ベルヌーイ分布 ($n = 1$ の二項分布)
- リンク関数: $\text{logit}(p) = \log \frac {p} {1 - p}$
何かの成否に対する何かの因子の影響、とか
風が吹けば桶屋が儲かる。
\[\begin{split} y_i &\sim \text{Bernoulli}(p_i) \\ &= \text{Binomial}(1,~p_i) \\ \text{logit}(p_i) &= \beta_0 + \beta_1 x_i \\ p_i &= \frac 1 {1 + e^{-(\beta_0 + \beta_1 x_i)}} \end{split}\]
ロジスティック関数↑
一般線形モデル (“化”無し) はGLMの一種
- 確率分布: 正規分布
- リンク関数: 恒等関数(なにもせずそのまま)
\[\begin{split} y_i &\sim \mathcal{N}(\mu_i,~\sigma^2) \\ \text{identity}(\mu_i) &= \beta_0 + \beta_1 x_i \end{split}\]
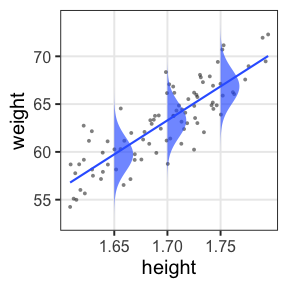
最小二乗法の直線あてはめと結果的に同じになる。
単回帰・重回帰と言ったとき一般線形モデルを前提とする人もいる。
分散分析 (Analysis of variance, ANOVA) as GLM
質的な説明変数を持つ正規分布・恒等リンクのGLM、と解釈可能。
指示変数 (0 or 1) に変換してから重回帰する。
| 天気 | → | $x_1$ ☀️ 晴れ | $x_2$ ☔️ 雨 |
|---|---|---|---|
| ☁️ くもり | 0 | 0 | |
| ☀️ 晴れ | 1 | 0 | |
| ☔️ 雨 | 0 | 1 |
\[\begin{split} y_i &= \mathcal{N}(\mu_i,\sigma^2) \\ \mu_i &= \beta_0 + \beta_1 x_{1i} + \beta_2 x_{2i} \end{split}\]
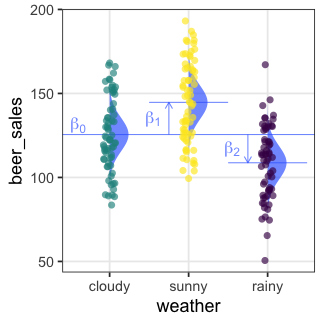
くもり☁️ $\beta_0$ を基準に、晴れの効果☀️ $\beta_1$ と雨の効果☔️ $\beta_2$ が求まる。
GLMなら確率分布・リンク関数を変えてもっと柔軟にモデリングできる。
共分散分析 (Analysis of covariance, ANCOVA) as GLM
質的変数と量的変数を両方含むGLM、と解釈可能。
正規分布・等分散・恒等リンクなどが仮定される。
| 天気 | → | $x_1$ ☀️ 晴れ | $x_2$ ☔️ 雨 |
|---|---|---|---|
| ☁️ くもり | 0 | 0 | |
| ☀️ 晴れ | 1 | 0 | |
| ☔️ 雨 | 0 | 1 |
\[\begin{split} y_i &= \mathcal{N}(\mu_i,\sigma^2) \\ \mu_i &= \beta_0 + \beta_1 x_{1i} + \beta_2 x_{2i} + \beta_3 x_{3i} \end{split}\]
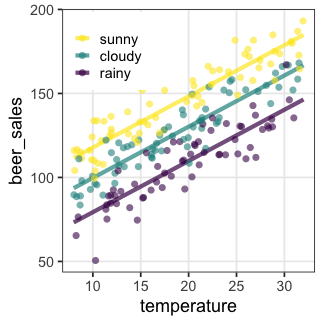
GLMなら確率分布・リンク関数を変えてもっと柔軟にモデリングできる。
一般化線形モデル(GLM)ふりかえり
確率分布・リンク関数を変えて柔軟にモデリングできる。
特定の組み合わせには名前がある。
| 名前 | 確率分布 | リンク関数 | 説明変数 |
|---|---|---|---|
| ポアソン回帰 | ポアソン分布 | log | |
| ロジスティック回帰 | 二項分布 | logit | |
| 一般線形回帰 | 正規分布 | 恒等 | |
| 分散分析 | 正規分布 | 恒等 | 質的変数 |
| 共分散分析 | 正規分布 | 恒等 | 質的変数+量的変数 |
リンク関数をもう少しだけ掘り下げたい。
リンク関数
統計モデリングにおいて「まっすぐ以外も表現できる」意味
- $\text{identity}(\mu_i)$
- $\mu_i = \beta_0 + \beta_1 x_{1i} + \beta_2 x_{2i} + \ldots$
- 説明変数の効果が足し算的に働く。
- $\log(\lambda_i)$
- $\lambda_i = e^{\beta_0 + \beta_1 x_{1i} + \beta_2 x_{2i} + \ldots} = e^{\beta_0} \times e^{\beta_1 x_{1i}} \times e^{\beta_2 x_{2i}} \times \ldots$
- 説明変数の効果が掛け算的に働く。
e.g., $\Delta x_1$ 増えると $e^{\beta_1 \Delta x_{1}}$ 倍になる - $\text{logit}(p_i)$
- $p_i = \frac 1 {1 + e^{-(\beta_0 + \beta_1 x_i + \ldots)}} $ (ロジスティック関数)
- 説明変数の効果が頭打ちになる。
e.g., $\lim_{x \to -\infty} p = 0;~\lim_{x \to \infty} p = 1$
ほかに probit, inverse, sqrt, etc.
データはひとつ、モデルはたくさん
どう選ぶ?
- メカニズム的に納得できるものを選ぶ
- ポアソン過程のカウントならポアソン分布、間隔ならガンマ分布
- n回中k回のように割合的なカウントなら二項分布
- データを可視化してみて、それっぽい形・性質のものを選ぶ
- 左右対称のひと山ならとりあえず正規分布
- 負の値を取らないならガンマ分布
- 直線的か、指数関数的か、頭打ちか、などなど
客観的な指標もほしい。
モデルの尤もらしさといえば…
尤度 (likelihood)
あるモデル$M$の下でそのデータ$D$が観察される確率:
$\text{Prob}(D \mid M)$
データ$D$を固定し、モデル$M$の関数とみなしたものが尤度関数:
$L(M \mid D)$
モデルの構造も固定してパラメータ$\theta$だけ動かす場合はこう書く:
$L(\theta \mid D)$ or $L(\theta)$
対数尤度 $\log L$ の形にしたほうがいろいろ便利。
各モデルで最適なパラメータを探して、比較:
$\log L^* (M_1) \text{ vs. } \log L^* (M_2) \text{ vs. } \log L^* (M_3) \ldots$
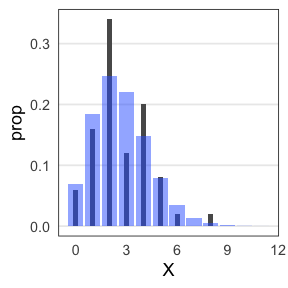
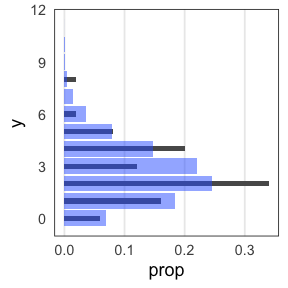
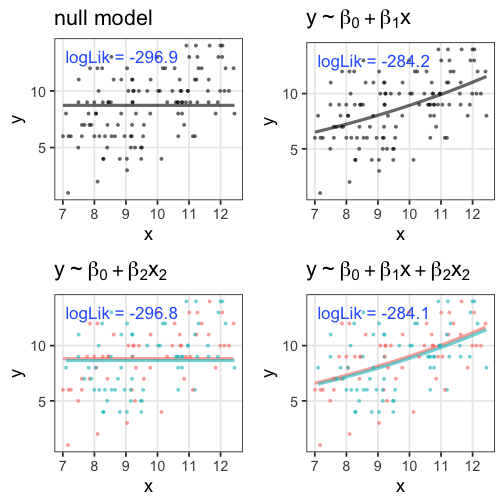
たしかに尤度はあてはまりの良さを表してそう
この場合は直線回帰よりもポアソン回帰が良さそう:
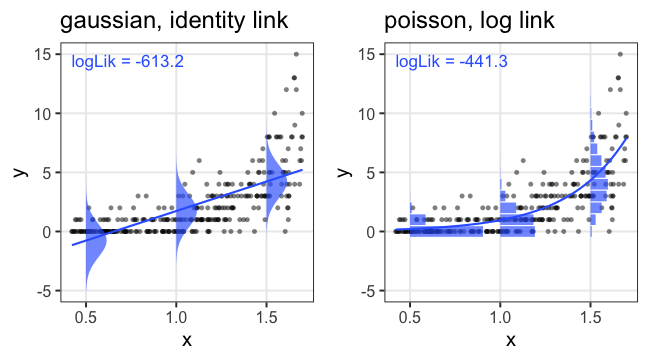
この調子で、より尤度の高いモデルを探していけばいいだろうか?
あてはまりが良ければいいってもんでもない
- 過剰適合 / 過学習 / overfitting
- パラメータを増やせば現データへの適合度・尤度を高くできるが、
予測・理解の役には立たなくなる。
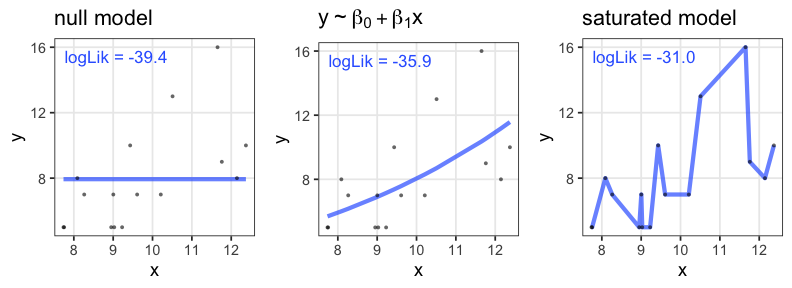
帰無モデル: 説明変数なし。切片のみ。
飽和モデル: データ点の数 ≤ パラメータの数。“データ読み上げ”的モデル
無駄な説明変数を加えても尤度は上がる
ある植物が作る種の数 $y$ は個体のサイズ $x$ に応じて増える。
観察時に着てた服の色 $x_2$ を追加すると尤度が上がる……?
AIC: 赤池情報量基準
\[\begin{split} \text{AIC} = -2 (\log L^* - k) = -2 \log L^* + 2k \end{split}\]
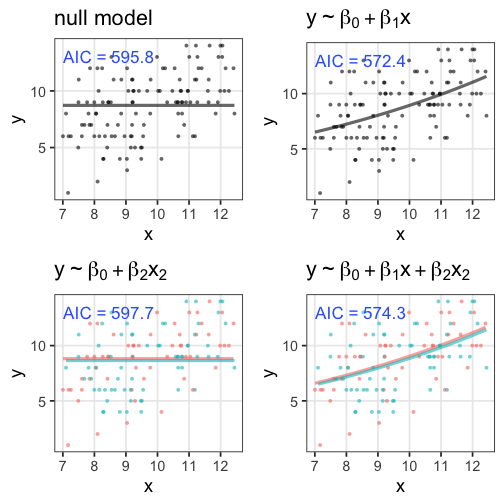
- AICが小さいほど予測精度の良いモデル。
- 尤度は上げたい。
- パラメータ数 $k$ が増えるとペナルティ。
- どのデータに対する当てはまりを目指すかという観点
- 「手元のデータ」に対する対数尤度は $\log L^*$
- 「真のメカニズムから出てくる未来のデータ」に対する
平均対数尤度の推定量は $(\log L^* - k)$
(Kullback–Leibler情報量を使って導出するらしい)
- 「手元のデータ」に対する対数尤度は $\log L^*$
無駄な説明変数の追加でAIC増加
ある植物が作る種の数 $y$ は個体のサイズ $x$ に応じて増える。
観察時に着てた服の色 $x_2$ を追加したモデルはAICが増加。
ほかの情報量基準
モデル選択の心構え
「正しい」ものを選べるわけではない。
予測・理解に useful なものを何らかの基準で選ぶだけ。
All models are wrong, but some are useful. — George E. P. Box

現実的な注意点・悩みどころ
- 多重共線性(multicollinearity):
- 説明変数同士が強い相関関係にある
- 変数変換:
- 気安くやるべきじゃないけど、対数変換などしばしば有用
- 割り算した値は危険
- 交互作用を入れると解釈が難しくなる。
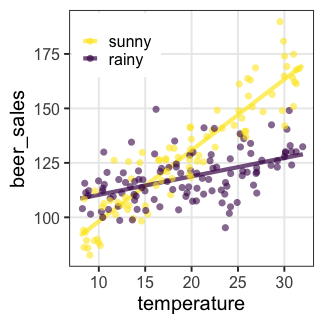
交互作用
ある説明変数の効果が、別の説明変数によって異なる。
e.g., ビール売上の温度依存性が天気によって異なる。
| 天気 | $x_1$ |
|---|---|
| ☀️ 晴れ | 1 |
| ☔️ 雨 | 0 |
\[\begin{split} y_i &= \mathcal{N}(\mu_i,\sigma^2) \\ \mu_i &= \beta_0 + \beta_1 x_{1i} + \beta_2 x_{2i} + \beta_{1,2} x_{1i} x_{2i} \end{split}\]
雨の日は $x_{1i} = 0$ のため $\beta_0,~\beta_2$ の項だけ。
晴れの日はそれに加えて $\beta_1,~\beta_{1,2}$ の項も。
解釈が一気に難しくなるのでむやみに使わない。
一般化線形モデル座学まとめ
- 何はともあれ散布図を描く
- 適切な確率分布・リンク関数・説明変数を考える
- パラメータを最尤推定する
- 尤度は「手元のデータへのあてはまり」
- モデルを比較するときは情報量基準を参考にする
penguinsデータセット

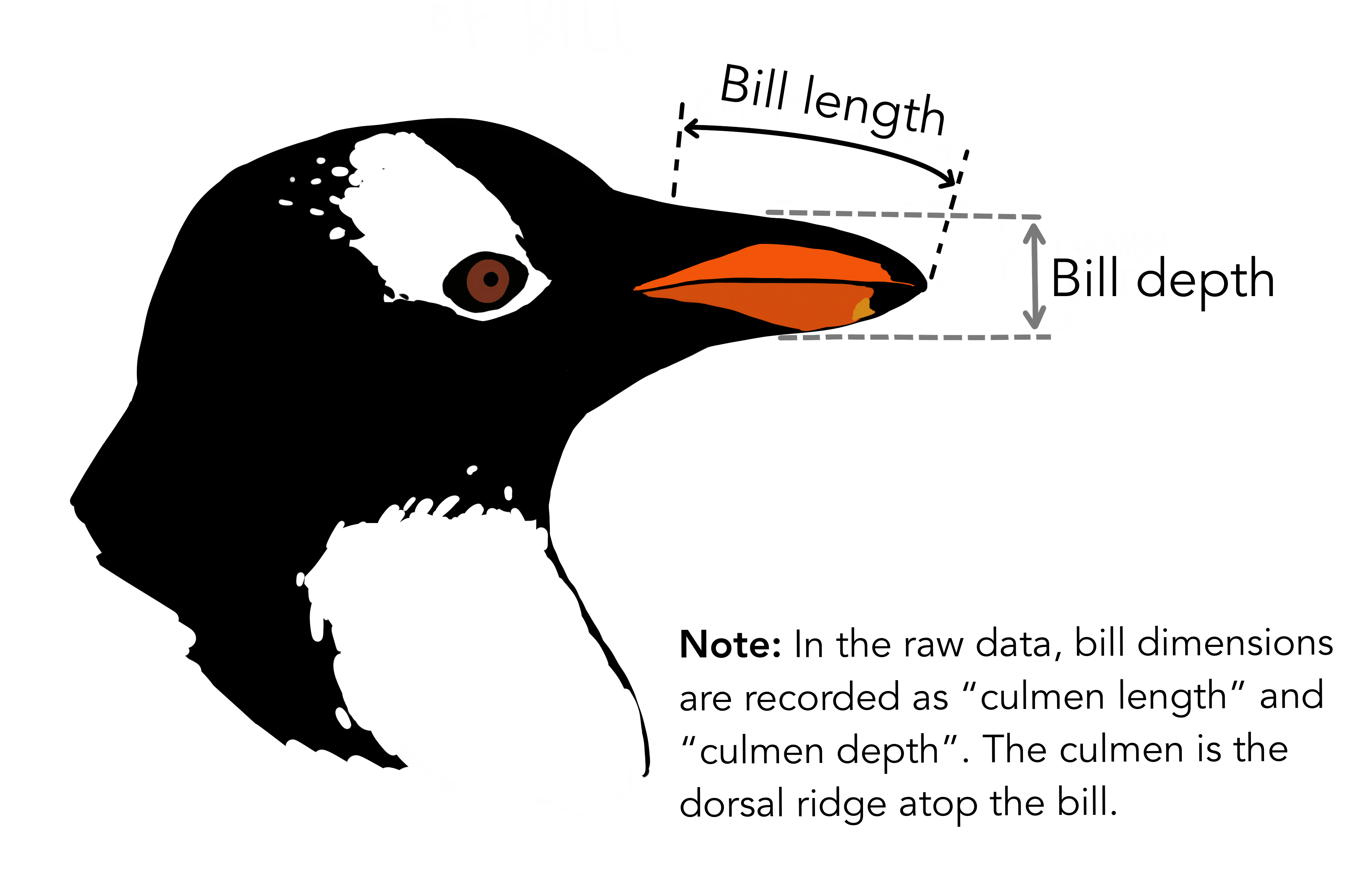
# install.packages("palmerpenguins")
library(palmerpenguins)
penguins_colors = c(Adelie = "darkorange", Chinstrap = "purple", Gentoo = "cyan4")
print(penguins)
penguinsデータセット
species island bill_length_mm bill_depth_mm flipper_length_mm body_mass_g sex year
<fct> <fct> <dbl> <dbl> <int> <int> <fct> <int>
1 Adelie Torgersen 39.1 18.7 181 3750 male 2007
2 Adelie Torgersen 39.5 17.4 186 3800 female 2007
3 Adelie Torgersen 40.3 18.0 195 3250 female 2007
4 Adelie Torgersen NA NA NA NA NA 2007
--
341 Chinstrap Dream 43.5 18.1 202 3400 female 2009
342 Chinstrap Dream 49.6 18.2 193 3775 male 2009
343 Chinstrap Dream 50.8 19.0 210 4100 male 2009
344 Chinstrap Dream 50.2 18.7 198 3775 female 2009
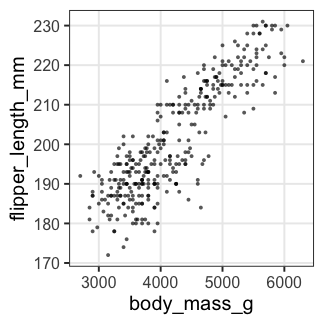
単回帰の練習: 1. まず作図
どうやら、重いペンギンほど翼長も長い。
p_penweight = ggplot(penguins) +
aes(body_mass_g, flipper_length_mm) +
geom_point(shape = 16, alpha = 0.66) +
theme_bw(base_size = 20) +
theme(panel.grid.minor = element_blank())
p_penweight
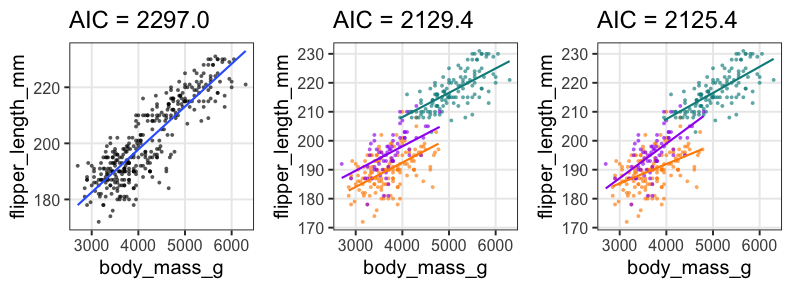
単回帰の練習: 2. モデル作成、フィッティング
fit1 = glm(flipper_length_mm ~ body_mass_g, data = penguins)
broom::tidy(fit1)
term estimate std.error statistic p.value
<chr> <dbl> <dbl> <dbl> <dbl>
1 (Intercept) 136.72955927 1.996835406 68.47312 5.712947e-201
2 body_mass_g 0.01527592 0.000466836 32.72223 4.370681e-107
broom::glance(fit1)
null.deviance df.null logLik AIC BIC deviance df.residual nobs
<dbl> <int> <dbl> <dbl> <dbl> <dbl> <int> <int>
1 67426.54 341 -1145.518 2297.035 2308.54 16250.3 340 342
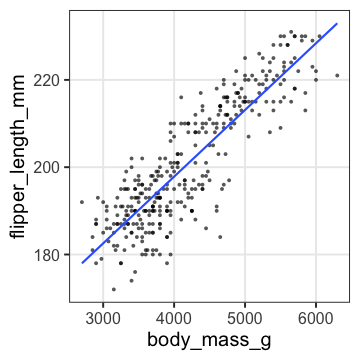
単回帰の練習: 3. フィッティング結果を作図
$y = 136.7 + 0.0153 x$
added1 = modelr::add_predictions(penguins, fit1)
p1 = p_penweight +
geom_line(aes(y = pred), data = added1, size = 1, color = "#3366ff")
p1
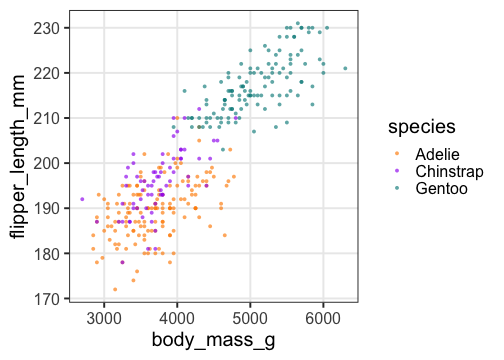
重回帰の練習: 1. まず作図
重いペンギンほど翼長も長い。翼長は種によっても違うかも。
p_penweight_color = p_penweight + aes(color = species) +
scale_color_manual(values = penguins_colors)
p_penweight_color
重回帰の練習: 2. モデル作成、フィッティング
Adelieを基準に、ChinstrapとGentooはそれより長め。
体重の効果は単回帰のときより小さい。
fit2 = glm(flipper_length_mm ~ body_mass_g + species, data = penguins)
broom::tidy(fit2)
term estimate std.error statistic p.value
<chr> <dbl> <dbl> <dbl> <dbl>
1 (Intercept) 1.588603e+02 2.3865766963 66.564071 2.450113e-196
2 body_mass_g 8.402113e-03 0.0006338976 13.254686 1.401600e-32
3 speciesChinstrap 5.597440e+00 0.7882166229 7.101398 7.334777e-12
4 speciesGentoo 1.567747e+01 1.0906590679 14.374308 6.800823e-37
broom::glance(fit2)
null.deviance df.null logLik AIC BIC deviance df.residual nobs
<dbl> <int> <dbl> <dbl> <dbl> <dbl> <int> <int>
1 67426.54 341 -1059.718 2129.437 2148.611 9839.073 338 342
重回帰の練習: 3. フィッティング結果を作図
added2 = modelr::add_predictions(penguins, fit2)
p2 = p_penweight_color +
geom_line(aes(y = pred), data = added2, size = 1)
p2
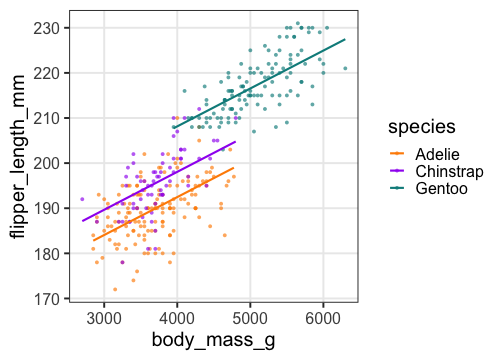
傾きも種によって違うかも。交互作用を入れてみたい。
交互作用の練習: モデル作成、フィッティング
Adelieを基準に、Chinstrapの傾きが結構違う。
切片の違いは解釈しにくくなった。
fit3 = glm(flipper_length_mm ~ body_mass_g * species, data = penguins)
broom::tidy(fit3)
term estimate std.error statistic p.value
<chr> <dbl> <dbl> <dbl> <dbl>
1 (Intercept) 165.244812649 3.5508916651 46.536146 1.561669e-148
2 body_mass_g 0.006676867 0.0009522935 7.011354 1.301783e-11
3 speciesChinstrap -13.863939075 7.3012647809 -1.898841 5.844186e-02
4 speciesGentoo 6.059375933 6.0508813200 1.001404 3.173522e-01
5 body_mass_g:speciesChinstrap 0.005228197 0.0019486293 2.683013 7.657147e-03
6 body_mass_g:speciesGentoo 0.002362269 0.0013525781 1.746494 8.163897e-02
broom::glance(fit3)
null.deviance df.null logLik AIC BIC deviance df.residual nobs
<dbl> <int> <dbl> <dbl> <dbl> <dbl> <int> <int>
1 67426.54 341 -1055.711 2125.422 2152.265 9611.166 336 342
交互作用の練習: フィッティング結果を作図
added3 = modelr::add_predictions(penguins, fit3)
p3 = p_penweight_color +
geom_line(aes(y = pred), data = added3, size = 1)
p3
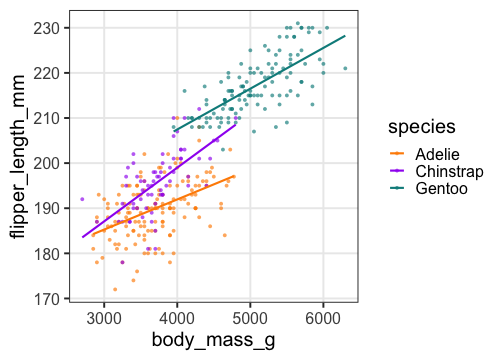
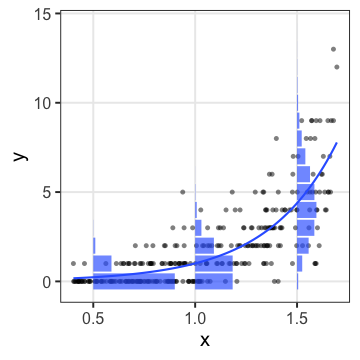
ここまでの3つのモデルでどれがいいか?
AICで選ぶなら交互作用入り重回帰が良さそう。
AIC(fit1, fit2, fit3)$AIC
GLMの練習
🔰クチバシの長さと深さで同じ解析をやってみよう。
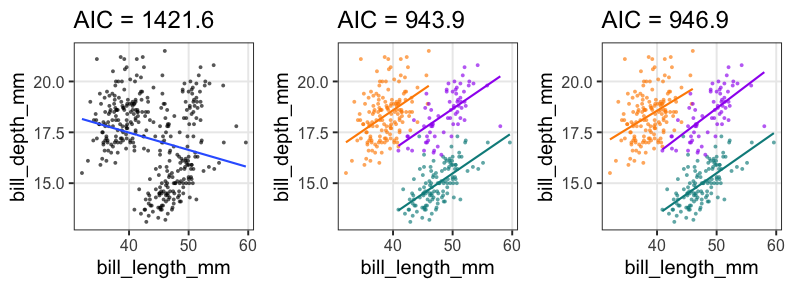
🔰余裕があったら性別や年なども説明変数に入れてみよう。
確率分布とリンク関数を明示的に指定したい
何も指定しない場合は正規分布・恒等リンクだった:
formula = flipper_length_mm ~ body_mass_g
fit1 = glm(formula, data = penguins)
fit1$family
Family: gaussian
Link function: identity
こう書いたのと同じ:
glm(formula, data = penguins, family = gaussian(link = identity))
利用可能な確率分布リンク関数は ?family などを参照。
🔰 4日目の課題1
- これまでに登場したデータセット、何でもいいのでGLMを適用してみよう。
- 確率分布とリンク関数を差し替えてAICの変化を確認しよう。
- 正規分布じゃないほうが当てはまりのいいデータを探してみよう。
4/18月曜はまず40分ほど班ごとに相談。
その後、各班の代表事例な発表してもらいます。
- データの出どころ、概要
- 読み込み、前処理、可視化、フィッティングのコード
- できあがりの図
ここ以降は余裕があれば進む
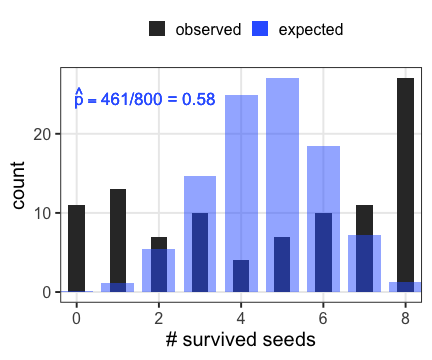
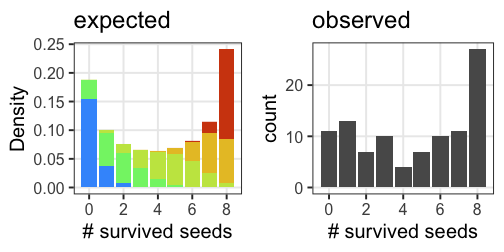
n個のうちy個生存。二項分布に従……わない!
植物100個体から8個ずつ種子を取って植えたら全体で半分ちょい発芽。
親1個体あたりの生存数はn=8の二項分布になるはずだけど、
極端な値(全部死亡、全部生存)が多かった。個体差?
個体差をモデルに組み込みたい
各個体の生存率$p_i$をそのままパラメータにすると過剰適合。
「パラメータ数 ≥ サンプルサイズ」の“データ読み上げ”モデル。
i.e., この個体は4個生き残って生存率0.5だね。次の個体は2個体だから……
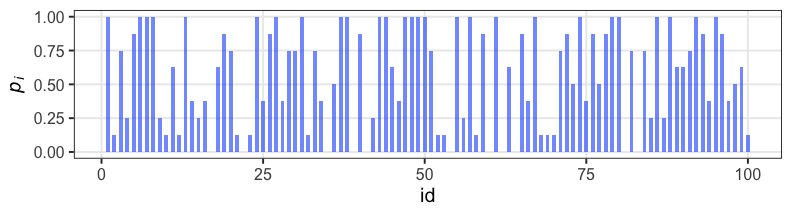
個体の生存能力をもっと少ないパラメータで表現できないか?
個体差をモデルに組み込みたい
各個体の生存率$p_i$が能力値$z_i$のシグモイド関数で決まると仮定。
その能力値は全個体共通の正規分布に従うと仮定:
$z_i \sim \mathcal{N}(\hat z, \sigma)$
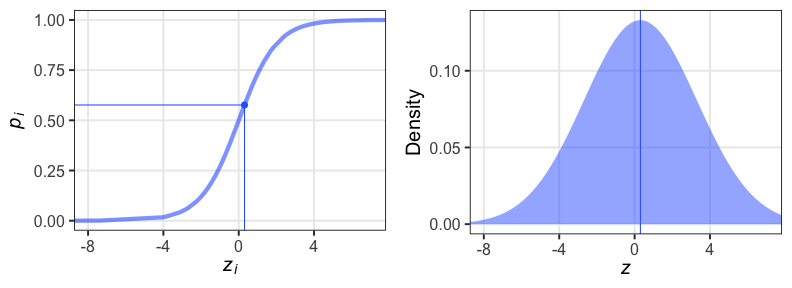
パラメータ2つで済む: 平均 $\hat z$, ばらつき $\sigma$ 。
前者は標本平均 $\hat p$ から求まるとして、後者どうする?
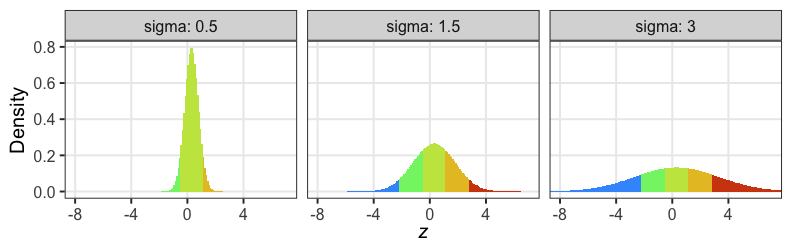
個体能力のばらつき $\sigma$ が大きいと両端が増える
普通の二項分布は個体差無し $\sigma = 0$ を仮定してるのと同じ。
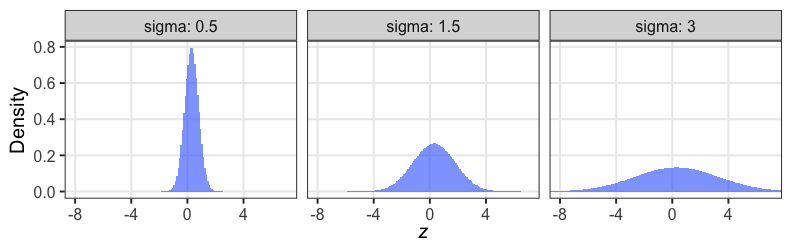
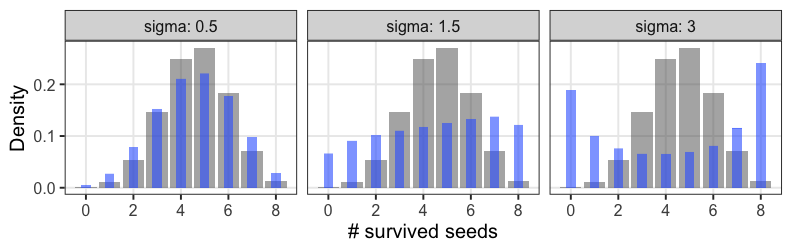
zの値で色分けしてみると想像しやすい
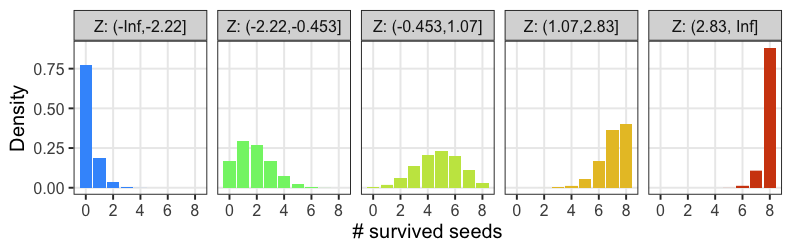
正規分布と二項分布の混ぜ合わせ……?
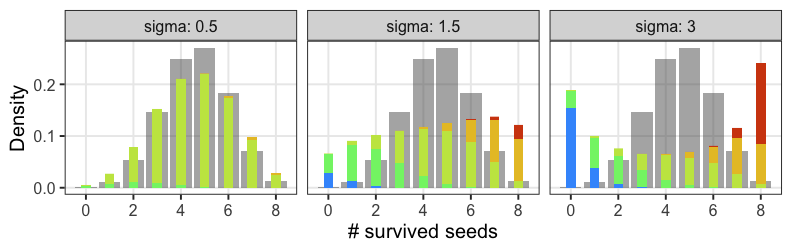
混合分布。ただの二項分布よりも良いあてはまり。
パラメータp(を決めるz)ごとに二項分布を作って、重み付けして足したもの。
一般化線形混合モデル GLMM
固定効果(fixed effects) のみ扱っていたGLMを拡張して、
変量効果(random effect) を混合したモデル。
「混合分布を使うモデル」という意味ではないらしい。
\[\begin{split} y_i &\sim \text{Binomial}(n,~p_i) \\ \text{logit}(p_i) &= \beta_0 + \beta_1 x_{1i} + \beta_2 x_{2i} + \ldots + z_{1i} + \ldots \\ z_{1i} &\sim \mathcal{N}(\mu_1,~\sigma_1) \end{split}\]
e.g.,
個体$i$の種子生存率$p_i$は、
(固定効果) 体サイズ$x_{1i}$と日当たり$x_{2i}$に依存し、
(変量効果) よくわからん個体差$z_{1i}$と植木鉢差$z_{2i}$もある。
固定効果にするか、変量効果にするか
推定したパラメータを予測に使うなら固定効果
- 予測に使えそうなので固定効果向き
- 観測・操作した連続値変数: 長さ、重さ、温度、etc.
- 観測・操作したカテゴリカル変数: 性別、投薬、etc.
- 予測に使えないので変量効果向き
- 観測・操作できなかった個体差:
たまたま集まってくれた学生15人 {A, B, C, …}。
Aさんの固定効果を推定できても、Zさんの予測には使えない。
- 観測・操作できなかった個体差:
- 観測・操作できなかったグループ差:
↑の学生をランダム5人ずつに分けたグループ {い、ろ、は}。
いグループの固定効果を推定できても、また集まることはない。
- 観測・操作できなかったグループ差:
どういうときに変量効果を考える必要があるか
データに擬似反復が含まれるとき。
ぜんぶ独立のつもりで解析すると推定が偏ったり誤ったり。
| 植木鉢 | 個体/植木鉢 | 種子/個体 | 疑似反復 | 推定不可 |
|---|---|---|---|---|
| 100個 | 1個体ずつ | 1個ずつ | – | 個体差・鉢差 |
| 25個 | 1個体ずつ | 4個ずつ | 個体 | 鉢差 |
| 20個 | 5個体ずつ | 1個ずつ | 植木鉢 | 個体差 |
| 5個 | 5個体ずつ | 4個ずつ | 植木鉢・個体 | – |
疑似反復あり
→ 観測できなかった個体差・場所差(変量効果)を推定可能
→ そのぶんを差し引いて固定効果を推定したい
GLMMの問題点・展望
- 最尤推定の計算が難しくなるので、あまり複雑にはできない
- ベイズ推定を使えばクリアできる
- GLMの拡張として理解はできても、実際に書くのは難しめ
- 階層ベイズモデルの一種として見るほうが便利
→ ここでGLMMの練習はせず、階層ベイズモデルに進む。
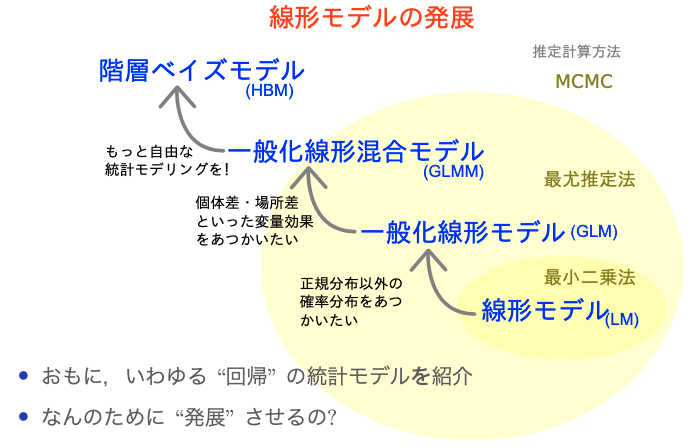
一般化線形(混合)モデルまとめ
- 何はともあれ作図して俯瞰
- GLMは統計モデリングの考え方の根幹
- 確率分布・リンク関数・説明変数
- 尤度・最尤法によるパラメータ推定
- 情報量基準などによるモデル選択
- GLMMは現実のデータ解析に向けた強化
- 疑似反復による変量効果を考慮
- 階層ベイズモデルとして扱うほうが楽
参考文献
- データ解析のための統計モデリング入門 久保拓弥 2012
- データ分析のための数理モデル入門 江崎貴裕 2020
- 分析者のためのデータ解釈学入門 江崎貴裕 2020
- 統計学を哲学する 大塚淳 2020
- 科学とモデル—シミュレーションの哲学 入門 Michael Weisberg 2017
(原著: Simulation and Similarity 2013)